Содержание:
 Железо — незаменимый микроэлемент почти для каждого живого организма. Так как железо легко принимает или отдает электроны, то свободное железо обладает высокой реакционной способностью и токсично. В биологических организмах его химическая активность ограничивается и направляется путем его ассоциации с простетическими группами и белками. Белки могут содержать железо как простетическую группу, виде железо-серных кластеров, гема, комплексов с остатками аминокислот в белках, в том числе с гистидином, глутаматом, аспартататом и тирозином.
Железо — незаменимый микроэлемент почти для каждого живого организма. Так как железо легко принимает или отдает электроны, то свободное железо обладает высокой реакционной способностью и токсично. В биологических организмах его химическая активность ограничивается и направляется путем его ассоциации с простетическими группами и белками. Белки могут содержать железо как простетическую группу, виде железо-серных кластеров, гема, комплексов с остатками аминокислот в белках, в том числе с гистидином, глутаматом, аспартататом и тирозином.
Железосодержащие белки, значание
- переносят или хранят кислорода (например, гемоглобин или миоглобин),
- являются ферментами,
- компонентами сигнальных путей,
- компонентами обмена ксенобиотиков — цитохром Р450
- антимикробных окислительно-восстановительных систем (например, цитохромы, рибонуклеотидредуктазы, синтаза оксида азота, NADPH оксидаза, миелопероксидаза);
- осуществляют транспорт или хранение железа (например, трансферрин , лактоферрин , ферритина ) .
- энергетический метаболизм,
- пластический метаболизм, включая синтез нуклеотидов,
- иммунная защита.
Хотя железо является одним из самых распространенных элементов в земной коре, дефицит железа — распространенное состояние для большинства организмов, живущих в условиях богатой кислородом Земли. Этот парадокс связывают к низкой растворимостью природных окисленных форм железа. Железо растительных продуктов у человека всасывается неэффективно, так как находится в форме нерастворимых комплексов, что находит свое отражение в высокой распространенности дефицита железа в популяциях людей, которые потребляют преимущественно вегетарианскую пищу. Гем железа из мяса, птицы и рыбы эффективно всасывается в кишечнике, но доступность мясных продуктов для человека в эволюции стала недавно и остается социально и географически ограниченной. Так как люди и другие млекопитающие эволюционировали в среде, где дефицит железа был обычным явлением, то для эффективного сохранения и внутренней рециркуляции железа сформировались тонко регулируемые механизмы. В современном благоустроенном мире при избытке железа в пище большинство людей способны ограничить его всасывание и избегать токсичности чрезмерного накопления железа.
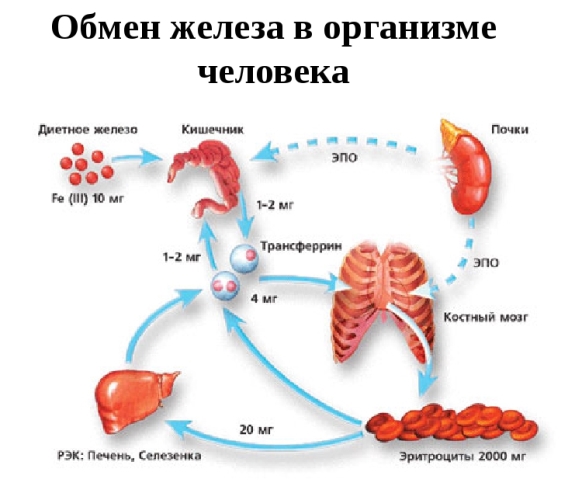
Средний взрослый человек содержит ~3-4 г железа, большинство из которого находится в гемоглобине эритроцитов (~2-3 г железа). Другие богатые железом ткани включают печень и селезенку, основные органы резерва железа. В этих органах железо хранится в макрофагах (фагоцитрующие клетки иммунной системы) и гепатоцитах (клетки печени), связанным со специфическим белком хранения железа, ферритином. Мышцы содержит железо в основном в миоглобине — белок накопления кислорода. Все клетки содержат железосодержащие белки, необходимые для производства энергии (АТФ), процессов синтеза нуклеиновых кислот, белков, жиров и углеводов и других важных функций.
Железо распределяется по тканям через плазму крови, которая содержит только 2-4 мг железа, связанного со специфическим белком переносчиком с трансферрином. За сутки трансферрин перемещает ~20-25 мг железа. Из всех клеток организма самую высокую концентрацию железа, ~ 1 мг / мл гематокрита — имеют эритроциты. Наибольшее количество железа плазмы — это железо состарившихся эритроцитов, которые поглощаются из кровотока макрофагами в селезенке и других органах и разрушаются ими. Так как продолжительность жизни эритроцитов человека является ~120 дней, то ~ 0,8 % (или ~15-25 мг ) железа всех эритроцитов должны быть переработаны каждый день. В свою очередь, из плазмы железо извлекается в основном для синтеза гемоглобина в предшественниках эритроцитов. Этот процесс регулируется эритропоэтином, уровень которого определяется степенью насыщенности тканей кислородом — оксигенацией. Несмотря на быстрый оборот и изменения в использовании железа, концентрация железа в плазме в целом стабильна, что свидетельствует о том, что освобождение утилизированного макрофагами железа в плазму регулируется.
Потери железа организмом составляют только 1-2 мг / день иобеспечиваются в основном через слущивание поверхностного эпителия кожи и слизистых. В норме эти потери уравновешиваются всасыванием железа из пищи в верхнем отделе двенадцатиперстной кишки. Запасы железа у большинства людей, потребляющих железо-адекватную пищу, устойчивы. Стабильный уровень железа в организме обеспечивается регуляторными механизмами.
Гомеостаз железа в организме: ткани и транспортеры
Ткани. После рождения в гомеостазе железа у человека принимают участие три ключевых типа клеток:
- энтероциты двенадцатиперстной кишки, которые всасывают железо пищи,
- макрофаги селезенки и печени, которые перерабатывают железо старых эритроцитов и других клеток,
- гепатоциты, которые хранят железо и могут отдавать его при необходимости.
Во время внутриутробного развития плацентарный синцитиотрофобласт переносит железа от матери к плоду. Выход железа из энтероцитов, макрофагов, гепатоцитов и синцитиотрофобласта в плазму регулирует гормон гепсидин, который псинтезируют гепатоциты (в том числе гепатоциты плода). Железо, не поступившее в плазму сохраняется в макрофагах, гепатоцитах и, возможно, в плаценте как запас. Железо, сохраняемое в энтероцитах быстро теряется из организма, потому что у человека эти клетки обновляются каждые 2-5 дней. Железо сброшенных энтероцитов попадают в фекалии. Таким образом, разделение железа между энтероцитами слизистой оболочки двенадцатиперстной и плазмы эффективно определяет содержание железа в организме.
Преобладающими формами железа в рационе человека являются гем, ферритин и трехвалентное железо в комплексе с другими макромолекулами. Кислая среда желудка и воздействие пищеварительных ферментов вызывает частичное высвобождение этих форм железа из перивариваемой пищи. Гем и не гемовое железо, видимому, поглощается отдельными механизмами, не исключено, что существует путь поглощения ферритина. О транспорте и метаболизме в энтероците гема и ферритина известно мало, перенос неорганического железа изучен детально.
Гемовое и негемовое железо в организме переносится через мембрану апикальной части энтероцитов, частичо запасается в виде ферритина, часть его выделяется в виде ферри иона через базолатеральную часть энтероцитов.
Аналогичная последовательность реакций имеет место в лизосомах макрофагов, когда они фагоцитируют стареющие эритроциты. В составе фаголизосом эритроциты и гемоглобин перевариваются, выделившийся гем окисляется гемоксигеназой-1 (HO-1), освобождая железо, которое либо сохраняется в цитоплазме в составе ферритина, либо экспортируется в плазму. Как в энтероцитах, так и макрофагах присутствуют два набора транспортеров и цитоплазматический белок хранения железа ферритина. Один набор транспортеров обеспечивает поглощения железа и поступление его в цитоплазму, а второй набор транспортеров передает железо из цитоплазмы в плазму крови.
Импорт железа
В апикальной части энтероцитов поглощение неорганического двухвалентного железа из просвета кишечника осуществляет транспортер двух валетных металлов DMT1. DMT1- это белок, проникающий через мембрану клеток (интегральный), экспрессируется на границе щеточной каемки двенадцатиперстной, вместе с трансферрином в эндосомах предшественников эритроцитов и других типах клеток, где DMT1 выполняет чрезвычайно важную роль транспорта железа, поставляемого трансферрином, в цитоплазму. DMT1 присутствует и в мембране макрофагов.
Эффективный транспорт железа по DMT1 зависит от концентрации двухвалентного железа и концентрации протонов на конттранспорт. Для транспорта железа через DMT1 необходимо восстановление трех валентного железа пищи в двух валентное железо. Эта реакция катализируется цитохромом B на поверхности энтероцитов двенадцатиперстной. Аскорбиновая кислота повышает активность редуктазы Dcytb, вероятно, выступая в качестве предпочтительного внутриклеточного донора электронов.
Импорт гема
В настоящее время известны два возможных переносчика гема в цитоплазму. Первый — белок-носитель гема 1 (HCP1), второй — транспортер гема млекопитающих. Транспортер гема млекопитающих локализуется в фаголизосомах макрофагов, предполагают, что он участвуют в транспорте гема, извлеченного из стареющих эритроцитов, извлечении железа гема.
Ферритин и цитоплазматическое хранение
Ферритин представляет собой сферический гетерополимерный белок, состоящий из 24 субъединиц тяжелого (Н) или легкого (L) типа. Относится к системному гомеостазу железа, цитоплазматический ферритин может хранить большое количество железа. H-субъединицы ферритина функционируют как феррооксидаза, окисляющая цитоплазматическое Fe2+ до Fe3+ — формы хранения.
Двухвалентного железа доставляется ферритина в цитоплазматических шаперонов, главным образом, поли (RC) связывающий белок 1 (PCBP1) (223). Выход железа из ферритина может происходить через закрытых порах или аутофагии и деградации лизосом ферритина (129, 240). Как железа переходит из ферритина до железа экспортеров не известно.
Растворимая, относительно бедная железом форма ферритина, находится в плазме крови. Эта форма — полимер из 24-субъединиц, представленными в основном L-ферритином, источником ферритина плазмы являются главным образом макрофаги. Концентрация ферритина в сыворотке в целом коррелируют с запасами железа, но не при всех физиологических и патологических состояниях. При патологических состояниях макрофаги могут содержать гораздо меньше или больше железа, чем запасы его в паренхиме ткани, или ситуации, в которых синтез ферритина обусловлен воспалением — ферритин — белок острой фазы.
Экспорт железа
Единственным известным млекопитающих железо экспортером железа у млекопитающих является белок ферропортин. Ферропортин содержится в мембранах клеток, участвующих в передаче железа в плазму — базолатеральные мембраны энтероцитов двенадцатиперстной , мембраны макрофагов, синусоидальные поверхности гепатоцитов, а базальная поверхность синцитиобласта плаценты. Ферропортин транспортирует из клетки Fe2+, а также Zn2+, но не двухвалентные Mn, Cu, или Cd; механизмы переноса не известны.
Экспорт железа из клеток зависит от состава семьи медьсодержащих феррооксидаз, в том числе церулоплазмина, гефестина и, возможно, циклопена, которые используют молекулярный кислород для окисления ферро в ферри, Церулоплазмин — содержащий медь белок экспрессируется в печени и сетчатке. В плазме содержится растворимая форма церулоплазмина. Гефестин и циклопен — трансмембранные белки, которые экспрессируется соответственно в энтероцитах и плаценте. Дефицит церулоплазмина препятствует как всасыванию железа в кишечнике, так и высвобождению железа из макрофагов и вызывает накопление железа в мозге и в гепатоцитах.
Экспортер гема
Митохондрии имеют решающее значение для метаболизма железа, будучи уникальной площадкой для синтеза гема и основным местомбиосинтеза железо-серных кластеров ([Fe-S]) . Точная регуляция транспорта железа в митохондрии имеет важное значение для биосинтеза, образования гемоглобина и сборки белка фелезо-серных кластеров в процессе синтеза эритроцитов.
Предполагается, что железо транспортируется в митохондрии с помощью митоферрина.
Идентифицированы три молекулы как возможные транспортеры гема из митохондрий, где осуществляется его синтез:
- белок резистентности к раку молочной железы
- ABC-митохондриальный эритроидный транспортер
- рецептор подгруппы С кошачьего вируса (FLVCR)
Внеклеточный транспорт железа
При нормальных обстоятельствах, трехвалентное железо экспортируется из клеток ферропортином и связывается с трансферрином -траспортером железа в плазме. Трасферрин может переносить до двух ионов трехвалентного железа, и доставить их в целевую ткань для поглощения с помощью рецептора трансферрина-1 (TfR1).
В дополнение к трасферрину, плазменные белки гемопексин и гаптоглобин связывают свободные гем и гемоглобин, соответственно, ограничивая их токсическое действие и способствуя утилизации в железо. Гемопексин и гаптоглобин играют важную гомеостатическую роль во время гемолитической стресса и заболеваний.
Гормональный контроль гомеостаза железа осуществляет гепсидин — гормон, синтезируемый печенью.